生物の進化と多様性(2009年5月5日)
生物の進化のプロセス
40億年ほど昔、灼熱の溶融状態だった地球表面が次第に冷やされて大陸と海洋が出現した頃、海洋の中にアミノ酸の離合集散により自己コピー機能を持った生物の原型が出現した。 そのコピーが不完全であった為、親とは少しずつ異なる生物体が種々出現し、それらのコピーのうち、厳しい環境に適応してより強い増殖力を持ったものが生き残り、弱いものは絶滅して行った。 コピー形式自体にも進化があった。 突然変異により色々なコピーシステムを持った原始生物が出現したが、現存する生物が殆どすべて行っている、DNA遺伝子を設計図として必要な蛋白質を合成しそれらを組み合わせて個体を構成するタイプの生物が生き残り、他のコピーシステムを採用した生物は淘汰され絶滅した。
進化の原動力は遺伝子の塩基配列の突然変異(コピーミス)だけではなく、異なる細胞同士の共生、同種の細胞がくっ付き合って起こる多細胞化などがあげられる。
すなわち、まずDNA遺伝子をコピーするとき僅かなコピーミスがある為、世代を重ねるにつれて新しい遺伝子配列を持った別の品種に多様化して行った。 それと平行して、初期の単細胞時代には、細胞と細胞との共生により新しい機能を持った品種の生物が出現する事もかなり起こった。 例えば、細胞の中にあり酸素を利用してエネルギーを発生させるミトコンドリアは、かって独立していた生物が他の生物の細胞に入って共生し、エネルギー効率の高い生物細胞を生み出したと考えられている。 その証拠に、細胞内の一器官で有りながら独自のDNAを持っている。 植物の細胞にある葉緑素も元は独立した生物であったが、他の細胞と共生して光合成する植物を生み出した、と推察される。
この他、単細胞生物が複数個集まって多細胞生物に変貌し、部分的な細胞の塊がそれぞれ機能を分担する器官へと組織分化した。 例えば昆虫の様に、餌を食べる口、消化器、呼吸器、運動する手足、飛揚する羽、簡単な神経系等を持つ多細胞生物に進化して行った。 人間の場合も、身体は機能を分担する色々な器官で構成され、細胞の総数は60兆個にもなる。
非常にシンプルなアメーバ的単細胞生物からDNAの突然変異、共生、多細胞化などのプロセスを繰り返し、厳しい自然淘汰を経て、約40億年の歳月を掛け、現存する数千万種を越える多様な生物へと進化して来た。 生物学者の推計によると、地球上には現在発見されている種の数倍以上の生物種が生存していると言う。
すなわち、生物の進化とは主としてコピーミスと自然淘汰を車の両輪とする非常にゆっくりとした長時間のプロセスであり、多様化とはその結果である。 これから更に10億年経つと、多様化は何処まで進むのであろうか。 種の数は経過時間に比例するのか、その何乗かに比例して指数関数的に増大するのか、考えて見よう。
進化の理論;中立仮説(木村資生編:分子進化学入門を参照、1968年木村氏がこの説を提唱)
進化の理論として現在最も有力な仮説である中立仮説によると、4種の塩基分子の二重螺旋配列であるDNAの各分子が置換する(例えば、A,T,G,Cの4塩基分子でAがTやGやCに置換わる)頻度は時間に比例する。その比例係数は遺伝子により異なり、生命維持に必須な大事な遺伝子の塩基分子の置換頻度はごく小さく、生命活動にあまり関係ない(生命維持にとって有利でも不利でもない中立的な変化で発生確率が一番高い)置換頻度は数倍以上大きくやはり経過時間に比例する。
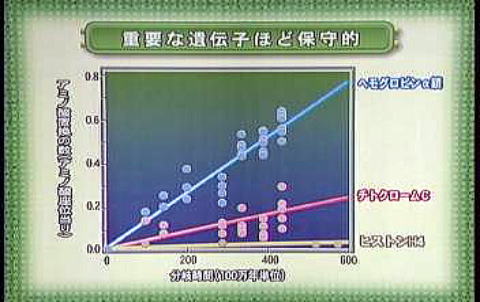
例えば、下図で示す様に、遺伝子の螺旋状塩基配列を更に螺旋状に巻上げて染色体の形状にする為に必要な蛋白質ヒストンは非常に重要な蛋白質であり、時間経過に対するアミノ酸の置換率は非常に低く保たれている。と言う事はアミノ酸配列を決定する遺伝子自体の塩基分子の置換速度も非常に低い事を意味する。 それに対し、赤血球のヘモグロビンα鎖のアミノ酸置換速度は非常に大きい。 これは、変化速度が大きくても生命活動を阻害しない、置換許容度の大きい蛋白質である事を示唆している。
別の例を示そう。例えば、ヒトとニワトリを比較した場合、インシュリンのA鎖では63個の塩基配列の内12個が違っており、B鎖では90個の塩基配列の内19個が違っている。
従って置換率λ=31/153=0.203となる。
一方、分子進化速度を定量的に表すには、長い時間![]() (例えば8000万年)だけ過去に分岐した2つの生物のある遺伝子の塩基配列を比較して、その間の進化の過程でK個の塩基置換が起きたとする。
(例えば8000万年)だけ過去に分岐した2つの生物のある遺伝子の塩基配列を比較して、その間の進化の過程でK個の塩基置換が起きたとする。
そうすると、進化速度![]() で表される。
で表される。
ここで、証明は省略するが、進化速度kと置換率λの関係はJukes-Cantorの式として次式で与えられる。
![]()
上のヒトとニワトリのケースでは、![]() =0.236±0.045となる。 ここで±0.045は推定値の誤差分散である。 (
=0.236±0.045となる。 ここで±0.045は推定値の誤差分散である。 (![]() は確率分布の平均であり、分散を伴う)
は確率分布の平均であり、分散を伴う)
なお上記の式では4つの塩基間の置換確率はどの組み合わせも全て等しいと仮定している。しかし現実の置換ではこの仮定は成立しない。 塩基TとC,AとGの置換と、塩基TとA、TとG、CとA、CとGの置換確率は異なる。 これらを補正すると
![]() =0.241±0.041となる。
=0.241±0.041となる。
ここで注意しなければならないのは、個体レベルでの突然変異の出現と、集団レベルでの突然変異体の固定とを、はっきり区別する事である。個体に表れた突然変異の大部分は少数の世代(例えば10世代)のうちに集団から失われてしまう。 そして幸運なほんの僅かなものだけが非常に長い時間かかって集団全体に広がる。これを突然変異の集団全体への“固定“と言う。また分子レベルの進化とは偶然発生したある個体内での突然変異ではなく、その変異が集団全体に普及・固定された状態を”進化“と言う。
自然淘汰に中立的な突然変異が固定する場合には、出現から固定までに平均![]() 世代かかる事が分かっている。ここに
世代かかる事が分かっている。ここに![]() は集団の有効な大きさで、ほぼ1世代あたりの総繁殖個体数と考えてよい。 例えば、
は集団の有効な大きさで、ほぼ1世代あたりの総繁殖個体数と考えてよい。 例えば、![]() 哺乳動物の集団で考えると、固定までの平均時間は
哺乳動物の集団で考えると、固定までの平均時間は
次に示すのは遺伝子の塩基置換ではなく、塩基置換が原因で生成されるアミノ酸の置換頻度を数種類の生物間で測定したものである。比較した物質は赤血球のα鎖と言う蛋白質のアミノ酸配列である。
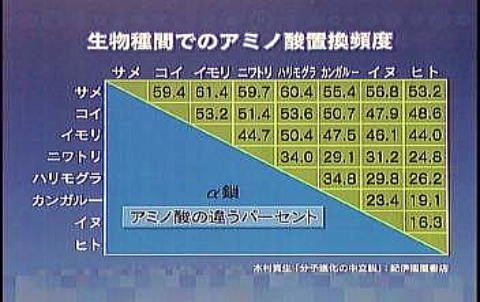
例えば、ヒトと犬では16.3%のアミノ酸が置換されているが、ヒトとサメでは53.2%の違いがある。置換速度は一定と考えられるので、この差異から両者が何万年前に同一祖先から分岐し、種の異なる生物になって行ったかが推測できる。
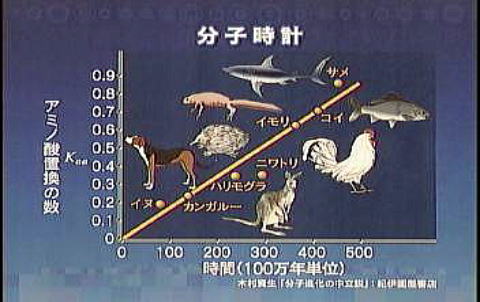
下のグラフは、ヒトとその他の動物が分岐した年代とアミノ酸置換率の関係を図示したものである。 両者の間にはほぼ直線的な関係がある事が分かる。 置換率によって分岐世代が分かるので、これは、進化のプロセス時間を計測できる一種の時計の役割をしており、“分子時計”と称される。
ある遺伝子の1世代当たり突然変異率を![]() とする。 また、集団内に出現した1個の突然変異遺伝子が究極的にその集団内に固定する確率(固定確率)を
とする。 また、集団内に出現した1個の突然変異遺伝子が究極的にその集団内に固定する確率(固定確率)を![]() とすると、
とすると、
進化速度![]()
が成立する。 その理由は、![]() 個体から成る集団中には毎世代
個体から成る集団中には毎世代![]() の新しい突然変異が出現し(
の新しい突然変異が出現し(![]() を2倍するのは、1個体を生じるには精子と卵子から対になる遺伝子を貰うからである)、その内の
を2倍するのは、1個体を生じるには精子と卵子から対になる遺伝子を貰うからである)、その内の![]() の割合のものが集団内に固定するからである。
の割合のものが集団内に固定するからである。
突然変異の大部分は自然淘汰の観点から見て中立的なものであるから、各個体に生じた突然変異には優劣はなく、ある個体の突然変異が集団全体に固定する確率はその集団の個体数分の1である。 すなわち、![]()
これを上式に代入すると、![]() すなわち、中立的突然変異に対しては、進化速度は遺伝子当たり毎世代当たりの突然変異率に等しいと結論される。
すなわち、中立的突然変異に対しては、進化速度は遺伝子当たり毎世代当たりの突然変異率に等しいと結論される。
進化速度![]() の実測値を例示すると次の様になる。
の実測値を例示すると次の様になる。
核 DNA遺伝子 ![]() 年
年
ミトコンドリアDNA遺伝子 ![]() 年
年
葉緑素DNA遺伝子 ![]() 年
年
インフルエンザ・ウイルス遺伝子 ![]() 年
年
種類の異なる生物の同義遺伝子(同じ働きをする遺伝子)を比較して塩基配列の違う箇所を数え、それをその遺伝子の全塩基数で割る。 例えばミトコンドリアの遺伝子を比較して、その数値が![]() と出れば、その数を進化速度
と出れば、その数を進化速度![]() で割り算すれば、それらの生物が共通祖先から分岐した年代が計算できる。 この場合は
で割り算すれば、それらの生物が共通祖先から分岐した年代が計算できる。 この場合は![]() 年、すなわち50万年昔に分岐した事が分かる。上記のアミノ酸置換率を分子時計と言う様に、塩基配列の進化速度も”分子時計“として利用できる事が分かる。
年、すなわち50万年昔に分岐した事が分かる。上記のアミノ酸置換率を分子時計と言う様に、塩基配列の進化速度も”分子時計“として利用できる事が分かる。
さて、ここで、生物が誕生して以来今日まで多種類に多様化して来た過程を中立論の理論を用いて定量的に推定して見よう。
ある生物集団の個体数を100,000とすると、突然変異が固定するまでの世代数は400,000世代となり、1世代10年とすると、4,000,000年で突然変異が集団内に固定し、新種が誕生する。 最初一種類だった生物が400万年毎に2種類に分岐したとすると、1億年経つと、生物種の数![]() となり、約3000万種に多様化する計算になる。 その内半数が絶滅したとすれば、残存種数は1500万となる。 現存する生物が30億年この様な進化をして来たとすると、4億5000万種の生物が現存する事になる。
となり、約3000万種に多様化する計算になる。 その内半数が絶滅したとすれば、残存種数は1500万となる。 現存する生物が30億年この様な進化をして来たとすると、4億5000万種の生物が現存する事になる。
化石とその地層を調べると、生物種の増加は指数関数的ではなく、何回も絶滅の危機を経過し、急激に減少したり、その後爆発的に多様化したりして現在に及んでいる。 従って本計算のような一様な条件を設定する事自体あまり意味が無いが、参考にはなる。